Diagram of muscle spindle
Image of muscle spindle
Muscle Spindles
Muscle spindles are found scattered throughout muscles and run in parallel with the main muscle fibres. They are important ‘proprioceptors’, i.e receptors that send sensory information to the CNS, that it uses to both to construct awareness of body position & movement and to enable the somatic nervous system to execute properly regulated movement programmes. The other main sensory organs contributing to propriception are Golgi tendon organs, joint receptors and pressure receptors (e.g. those in the sole of the foot).
The importance of proprioception is shown by the devastating effects of loss of the primary sensory neurons which subserve this sense. In the (fortunately very rare) cases of adults who lose their proprioceptive neurons, the result is effectively paralysis. Even though the skeletal muscles are all innervated normally by healthy motoneurons which in turn are controlled by undamaged upper motor neurons, the patient is unable to carry out volitional movements. This is because the CNS pathways which regulate motor programmes have been deprived of all the afferent information that they would normally receive about what the muscles are doing, and are consequently unable to activate the motor neurons.
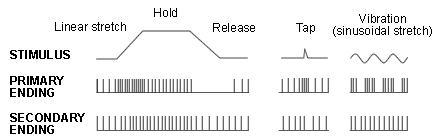
The spindle senses muscle length and changes in length. It has sensory nerve terminals whose discharge rate increases as the sensory ending is stretched. This nerve terminal is known as the Annulospiral ending, so named because it is composed of a set of rings in a spiral configuration.
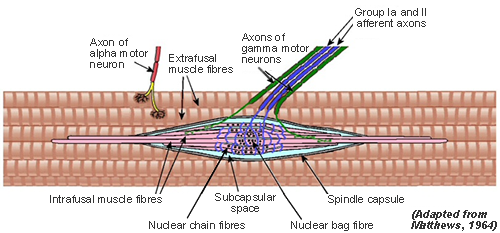
These terminals (shown in blue on the diagram) are wrapped around specialised muscle fibres that belongs to the muscle spindle (Intrafusal Fibres - see next tab) and are quite separate from the fibres that make up the bulk of the muscle (Extrafusal Fibres).
Traverse sections through muscle [Opens in new window]
Click on the Intrafusal Fibre tab above to continue.
Diagram of muscle spindle
Types of intrafusal fibre
There are two main types of intrafusal fibre: Nuclear bag and Nuclear chain.
Nuclear bag fibre
Nuclear bag fibre is so called because there is a bunch of about 100 nuclei in the central (or equatorial) region underlying the sensory nerve ending, which is of group Ia, the fastest-conducting found in the body
Another important feature is a motor supply to the intrafusal muscle (shown in green on the diagram). In this region on either side of the central area the intrafusal fibres are able to contract if their motor supply is active. The motor supply comes via efferent fibres that usually fall into the gamma classification of diameters. They are often (and better) referred to as Fusimotor fibres.
Two things can, in principle, cause the annulospiral ending to be stretched and so increase its discharge:
- A stretch of the muscle as a whole will stretch the spindles within it , and thus the sensory endings.
- Fusimotor activity will cause contraction of the intrafusal fibres below the fusimotor nerve terminals either side of the central region. This will result in stretch of the central sensory region .
Nuclear chain fibre
Nuclear chain fibres also have annulospiral sensory endings in the central region (where the nuclei are in line). This sensory nerve is of group Ia.
Further out we see that there are other sensory endings, more closely associated with the chain fibres. These fall into the slower group II division of sensory nerves and are referred to as Secondary endings in contrast to the centrally located Primary endings.
Summary
The two types of intrafusal fibre, (bag and chain) have different mechanical properties, and respond differently to their largely separate fusimotor fibres. They also differ in respect to their sensory endings. Consequently, the information relayed to the CNS by the spindle via group Ia and group II sensory endings is different.
Click on the Sensory Innervation tab above to continue.
Each spindle has one primary sensory ending, with spiral or annular (ring) terminations of a Group Ia afferent nerve which encircle all 3 types of intrafusal fibre.
There is also another kind of afferent, group II. Some spindles have none of these secondary sensory endings at all. Others have up to 5 (2 on one side 3 on the other side of the primary ending). Commonly there is one primary (P) with one secondary ending adjacent. Secondary endings are irregular spirals around chain fibres. Secondary endings vary in the contacts they make with bag fibres.
All 3 intrafusal fibres contribute to Group Ia output from the spindle (= input to the central nervous system).
Mainly chain fibres contribute to Group II output from the spindle (= input to the central nervous system).
In simple terms, the Ia afferents respond partly to muscle length, but respond more powerfully to changes in length. The group II afferents are much better at registering length alone.
Ia afferents can powerfully excite the alpha motoneurones of the muscle containing the spindle. This is the basis of the classical stretch reflex in which extension of the muscle (and thus its spindles) cause a reflex contraction. (see ‘Functional aspects of muscle spindles’ for more about this reflex).
The spindle can therefore register muscle length and velocity. Furthermore the sensitivity to both length and velocity can be altered by the CNS via activity in the fusimotor system, the static gamma system controlling length sensitivity and the dynamic gamma system controlling velocity sensitivity.
Click on the Motor Innervaton tab above to continue.
Two types of fusimotor innervation from gamma motoneurones in the spinal cord: dynamic and static (originally named on the basis of their effects of Ia afferent discharge during muscle stretch).
Dynamic gamma axons innervate dynamic bag fibres at one or both poles. Together they constitute the 'dynamic intrafusal system'.
The static bag fibres are innervated by static gamma axons.
Chain fibres have static gamma motor innervation.
For further details about motor innervation, see fusimotor effects on intrafusal fibres.
Select another item from the menu on left to continue.